Fishing for gene function – endocrine modelling in the zebrafish
- Department of Veterinary Basic Sciences, Royal Veterinary College, Royal College Street, London NW1 0TU, UK
- (Requests for offprints should be addressed to I M McGonnell; Email: imcgonnell{at}rvc.ac.uk)
Abstract
The use of zebrafish (Danio rerio) in scientific research is growing rapidly. It initially became popular as a model of vertebrate development because zebrafish embryos develop rapidly and are transparent. In the past 5 years, the sequencing of the zebrafish genome has increased the profile of zebrafish research even further, expanding into other areas such as pharmacology, cancer research and drug discovery. The use of zebrafish in endocrine research has mainly been confined to the study of the development of endocrine organs. However, it is likely to be a useful model in other areas of endocrinology, as there are a wide variety of both forward and reverse genetic techniques that can be employed in the zebrafish to decipher gene function in disease states. In this review, we compare the endocrine system of the zebrafish to mouse and human, demonstrating that the systems are sufficiently similar for zebrafish to be employed as a model for endocrine research. We subsequently review the repertoire of genetic techniques commonly employed in the zebrafish model to understand gene function in vertebrate development and disease. We anticipate that the use of these techniques will make the zebrafish a prominent model in endocrine research in the coming years.
Introduction
The zebrafish (Danio rerio), a member of the Cyprinidae group of teleosts, is a tropical freshwater fish, native to India and Burma. It has become a popular laboratory animal over the past 10 years and has some particular advantages for scientific research. Zebrafish are small – the adult grows to 50 mm in length (Fig. 1C⇓) – and can be kept in relatively high density. They are very easy to maintain under laboratory conditions, see Westerfield (2000) for more information on husbandry, which can also be accessed via ZFIN homepage http://zfin.org/cgi-bin/webdriver?MIval=aa-ZDB_home.apg. Their life span is approximately 3 years, but they have short generation times (3 months) and are reproductively capable for about 1.5 years. Each female can easily lay a clutch of 100–200 eggs weekly, which are externally fertilised and develop synchronously. They develop very rapidly (Fig. 1A and B⇓) and are free-feeding larvae within 5 days of fertilisation. The major application of the zebrafish in scientific research has been in developmental biology; this is due not only to the ease with which the embryos are obtained but also to the fact that the developing zebrafish embryo is transparent, greatly aiding observation of developmental processes and live imaging.
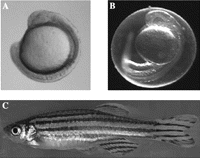
Developing and adult zebrafish. (A) Zebrafish embryo at 15 hpf. The embryo is curled around the yolk sac. The protective membrane chorion has been removed to aid clarity. Even at this early stage, the embryo has formed a neural tube and eyes as well as approximately 12 somites. (B) Zebrafish embryo at 24 hpf. This embryo has an intact chorion. Embryos at this stage undergo spontaneous movement of the tail. This image indicates the optical transparency of the zebrafish embryo. (C) Adult male zebrafish, 1 year. Magnification A and B, ×6; C, ×1.
Their popularity is now extending into other diverse areas of research such as physiology (Jagadeeswaran et al. 2005), cancer (Berghmans et al. 2005), drug discovery (Zon & Peterson 2005) and toxicology (Hill et al. 2005). This is in part due to the zebrafish genome-sequencing project, started in 2001, which has raised its profile even further as a genetic model. This project is updated daily, and can be accessed and searched via www.ensembl.org/Danio_rerio/index.html (see also ZFIN homepage for excellent genome resources and mutant database). Teleosts are thought to have undergone a whole genome duplication event during the early evolution of this group of fish, and thus zebrafish are tetraploid (reviewed in Taylor et al. 2001). There is abundant evidence that there have been some secondary losses of duplicate genes, possibly due to functional redundancy. However, there are also retained duplicate genes, some of which have the ancestral function divided between the two copies (subfunctionalisation) and some with new functions (neofunctionalisation) (reviewed in Wagner 1998; see also McClintock et al. 2001). Thus, the zebrafish affords a model to examine not only gene function but also the acquisition of new function through sequence change.
Endocrine disrupters
The main application of zebrafish in endocrine research until now has been in the investigation of the activity of endocrine disrupter chemicals (EDCs), particularly those that interfere with the function of the steroid hormones oestrogen and androgen (Ankley & Johnson 2004), which are known to play significant roles in development of gonads, maturation of germ cells and maintenance of reproductive cycles in all vertebrates (see below). Zebrafish are one of a group of small fish species that can be kept in the laboratory, are easily exposed to EDCs in tank water at different stages in the life cycle, and exhibit measurable sensitivity to EDCs, including sexual dimorphism (Van den Belt et al. 2001, Orn et al. 2003, Maack & Segner 2004). Aquatic species are particularly vulnerable to EDC, as many of these chemicals are water contaminants. However, small fish, such as zebrafish, may be good models for the effects of EDCs in mammals (Ankley & Johnson 2004).
Endocrine function in the zebrafish
There is a vast literature devoted to the endocrine system of fish due to the obvious scientific importance and economic benefit of understanding and manipulating this system. However, for fish to be a relevant model for the study of endocrine systems of other vertebrates (including man), they must be significantly similar in function. The following is a brief review of current knowledge of the development and basic functions of the fish endocrine system, highlighting the similarities and differences between that and the mammalian model. For more comprehensive information, we recommend the reviews quoted in each section below.
The hypothalamus
The hypothalamus acts as an endocrine gland in parallel with the pituitary gland. It develops from the ventral diencephalic region of the developing forebrain. Relatively few studies have investigated hypothalamic development in fish; however, a study by Mathieu et al.(2002) demonstrated that the prechordal plate plays a major role in hypothalamic development in the zebrafish. In contrast, the anterior endoderm plays this role in mammals.
The role of the hypothalamus can be divided broadly into two. The first is to release oxytocin and vasopressin into the bloodstream via the posterior pituitary. In fish, these hormones are called isotocin and vasotocin respectively. The second is to produce releasing factors (at least six), which in turn provoke the anterior pituitary to release hormones into the bloodstream. As modern teleosts lack a portal system, this is achieved through projecting axons that release these factors. Of these, gonadotrophin-releasing factor (GnRH) has been particularly well studied in fish due to the economic and ecological requirements for optimal breeding. There are thought to be three genes coding for GnRH isoforms in teleosts; however, zebrafish appear to only have two (Sherwood & Wu 2005). The majority of mammals also have two GnRH genes (Morgan & Millar 2004). As in mammals, they induce release of luteinising hormone (LH) and follicle-stimulating hormone (FSH) at different stages of maturation (reviewed in Ando & Urano 2005). In fish, as in other non-mammalian vertebrates, corticotrophin-releasing factor (CRF) stimulates not only corticotrophin release from the pituitary but also thyrotrophin release (Seasholtz et al. 2002). Thyrotrophin-releasing hormone (TRH) regulates both thyrotrophin and prolactin (PRL) release from the pituitary in mammals. It has been shown also to control growth hormone (GH) synthesis (Kagabu et al. 1998) and adrenocorticotrophic hormone (ACTH) secretion (Rotllant et al. 2000, 2001) in teleosts. In fish, growth hormone releasing-hormone (GHRH) and pituitary adenylate cyclase-activating polypeptide (PACAP) are coded for by one gene. In contrast, in mammals, they are coded for by two separate genes. Zebrafish PACAP sequence is highly conserved in mammals, but GHRH is much less conserved (Fradinger & Sherwood 2000). Zebrafish have duplicates, GRF-PACAP 1 (Fradinger & Sherwood 2000) and GRF-PACAP 2, the latter of which plays a role in oocyte maturation (Wang et al. 2003). Teleost PRL-releasing peptide (PrRP) has relatively low sequence homology to the human, but it is expressed in the hypothalamus and releases PRL from the pituitary. PrRP may also act to reduce levels of GH release, as seen in rats (Sakamoto et al. 2003).
In general, teleost hypothalamic hormones appear to have similar activities to mammalian equivalents. However, the additional roles found in some studies may point to a more complex system in the fish.
The pituitary
To date, the majority of work performed to elucidate developmental and functional aspects of the anterior pituitary has relied on mouse models and a range of human genetic abnormalities. Through these studies, an increasingly comprehensive plan of anterior pituitary development has been created, which involves considerable transcriptional regulation and signalling gradients of growth factors, leading to the spatial and temporal regulation of the endocrine lineages (for reviews, see Burgess et al. 2002, Scully & Rosenfeld 2002, Zhu et al. 2005). However, a reverse genetics approach to delineate the role of these transcription factors and signalling molecules in mice is often compromised by the early lethality of these models in utero, making phenotypic analysis almost impossible. The flexibility that the zebrafish offers in analysis of gene function makes it an attractive primary model for determining the process of pituitary organogenesis. For this reason, pituitary development is well studied in the zebrafish. The mature zebrafish anterior pituitary comprises the same five endocrine lineages expressed in the mammalian pituitary, although the sequence of their development is altered. Lactotrophs are the last endocrine lineage cell type to be expressed in mammals, yet they are the first lineage to differentiate in zebrafish (Herzog et al. 2003). In addition, there are key differences to the mammalian anterior pituitary in the morphogenesis and patterning within the zebrafish adenohypophysis. Specifically, whereas the mammalian anterior pituitary is derived from an invagination of the oral ectoderm, Rathke’s pouch, no such invagination occurs in zebrafish; instead, the anterior pituitary remains in a subepithelial position (Herzog et al. 2003). An additional structural difference is that the endocrine lineages are aligned anteroposteriorly (Herzog et al. 2003, 2004b) in contrast to dorsoventrally in the mammalian adenohypophysis (Dasen & Rosenfeld 1999).
Although obvious differences between mammalian and fish pituitary organogenesis exist, patterning seems well conserved. In mice, models have delineated a critical role for differentiating growth factors such as fibroblast growth factors (FGFs), bone morphogenetic proteins (BMPs) and the signalling molecule Sonic hedgehog (shh) (Dasen & Rosenfeld 1999, Sbrogna et al. 2003, Zhu et al. 2005). These same molecules are also critical to anterior pituitary development in zebrafish. For example, loss of Fgf3 function severely impairs zebrafish anterior pituitary development, resulting in loss of expression of pituitary hormones (Herzog et al. 2004a). Of further interest is the commonality between mammalian pituitary endocrine disorders and observed pituitary phenotypes in zebrafish mutants. For example, mutations in Pit1, which cause dwarfism in both mice (Li et al. 1990) and man (Pfaffle et al. 1992, Radovick et al. 1992, Tatsumi et al. 1992), are phenocopied in zebrafish Pit1 mutants, particularly in their lack of somatotrophs, lactotrophs and thyrotrophs (Li et al. 1990, Nica et al. 2004). However, consistent with the early appearance of lactotrophs, zebrafish Pit1 is also necessary for the expression of pituitary PRL.
Hormones performing the same or similar functions to mammalian pituitary hormones are expressed in the relevant endocrine lineages in zebrafish. In common with other fish, D. rerio express somatolactin (SL) as well as GH and PRL. Two isoforms, SLα and SLβ, have recently been cloned, SLα being more conserved with the majority of fish SLα genes, while SLβ shares homology with the more divergent goldfish SL (Zhu et al. 2004). PRL expression occurs earlier in zebrafish than mammals, where lactotrophs are the last endocrine lineage to develop. This suggests alternative roles for PRL in D. rerio, and this is supported by the fact that most pit1 mutant zebrafish die at 7–8 days post-fertilisation (dpf) (Nica et al. 2004), whereas mouse and human pit1 mutants are viable. SL has potential roles in regulating calcium metabolism, the stress response and acid–base balance in fish (Kakizawa et al. 1993), whereas PRL may perform important regulatory roles in freshwater adaptation by regulating cell proliferation and apoptosis (Sakamoto et al. 2005).
The zebrafish tshβ homologue is detected by 42 hpf, immediately before gh expression (Herzog et al. 2003). Developmentally, the onset of expression is reversed in zebrafish compared with their mammalian counterparts. The localisation of tshβ- and gh-expressing cells reveals that the majority of somatotrophs and thyrotrophs reside within the middle of the three regions that make up the mature anterior pituitary in zebrafish. The thyrotroph population are particularly susceptible to the effects of pit1 mutations, in that all thyrotrophs are lost (Herzog et al. 2004b), whereas only caudomedial thyrotrophs are lost in mouse models (Lin et al. 1993). This, perhaps, reflects the difference in when thyrotroph lineage cells arise in mice and zebrafish.
At least two POMC genes are expressed in zebrafish, which encode for ACTH, lipotrophin (γ-LPH), melanocyte stimulating hormone (β-MSH) and β-endorphin (Gonzalez-Nunez et al. 2003, Herzog et al. 2004b). In transgenic studies, corticotroph ontogeny was traced by driving expression of GFP with the zebrafish POMC promoter and revealed expression of POMC-GFP in bilateral groups anterior to the neural ridge midline at 18–20 h post-fertilisation (hpf) (Liu et al. 2003). These corticotroph lineage cells responded appropriately to dexamethasone treatment, which selectively expressed POMC-GFP by 2 dpf (Liu et al. 2003).
The cloning of zebrafish gonadotrophins has, until recently, been only partially completed. Nica et al.(2004) cloned a zebrafish homologue of the αgsu and found two distinct populations of cells; those that coexpressed pit1 (presumptive thyrotrophs) and those that did not (presumptive gonadotrophs). Most recently, the D. rerio genes for the gonadotrophins have been cloned (So et al. 2005). Two LH isoforms (lhβ1 and lhβ2), fshβ and GTHα sequences were cloned, and their distribution was restricted to the pituitary, testis and ovaries. Sequence homology of LH and αGSU with other fish species is reasonably high (~70%), but less conserved for FSH (37–64%) (So et al. 2005). In mammals, one of the key transcription factors responsible for the differentiation of gonadotroph lineage cells is steroidogenic factor-1 (SF-1), an ftz-f1 homologue. Zebrafish express two ftz-f1 homologues, zFF1 and ff1b, which are expressed at high levels in the pituitary and hypothalamus (von Hofsten et al. 2001). The zFF1 transcript also shares considerable homology with another closely related transcription factor, liver receptor homologue-1 (LRH-1), suggesting that the two ftz-f1 homologues in zebrafish can perform roles similar to both SF-1 and LRH-1. However, it remains to be elucidated as to whether LRH-1-like functions are required in the pituitary.
What is clear from this growing area of research is that, despite some notable exceptions, pituitary organogenesis and function in zebrafish are closely conserved with other vertebrates, making them an ideal system in which to dissect the role of specific pathways involved in pituitary development and function.
The parathyroid gland
The parathyroid gland detects levels of serum extracellular calcium via the calcium-sensing receptor. Low levels of calcium provoke the production and secretion of parathyroid hormone from the gland, which then acts to release calcium from stores in bone, reduce excretion from the kidney and increase absorption in the gut (Hogan et al. 2005). The parathyroid gland and parathyroid hormone are found in all tetrapods (land-dwelling vertebrates). However, fish do not possess a parathyroid gland, and there is a widespread belief that fish do not produce parathyroid hormone because of the different needs for calcium homeostasis in the aquatic environment. Calcium is abundant in water and is readily absorbed by fish, negating the need for storage. Despite this, fish do regulate plasma calcium levels, and in keeping with this, several recent studies have indicated that members of different groups of fish – zebrafish and puffer fish (both teleosts or bony fish) and dogfish (a Chondrichthyes or cartilaginous fish) –not only possess two parathyroid hormone genes (PTH 1 and 2) (Danks et al. 2003, Gensure et al. 2004, Hogan et al. 2004, Okabe & Graham 2004) but also produce active peptides (Danks et al. 2003, Gensure et al. 2004). Both the parathyroid hormone and the calcium sensing receptor gene are expressed not in a parathyroid gland but in the gills of adult fish (Okabe & Graham 2004). Gills are known to play a role in sensing/regulating calcium levels. Both the gills and the parathyroid gland are derived from the same embryonic tissue-pharyngeal pouch endoderm in the developing head, and require the Gcm2 gene for their development (Hogan et al. 2004, Okabe & Graham 2004). This suggests that, while fish do not possess a parathyroid ‘gland’, the gill is a homologous structure and fulfils the same function by producing parathyroid hormone, making the zebrafish a suitable model for the study of parathyroid hormone function.
The thyroid
The thyroid gland in the adult zebrafish is not a compact structure encapsulated in connective tissue, but rather a loose aggregation of follicles, close to the ventral aorta, distributed between the first gill and the heart (Wendl et al. 2002). Despite this, it develops from the same tissue as the thyroid in mammals – the endodermal tissue in the pharynx (Wendl et al. 2002) – indicating that they are homologous structures. Similarly, the genes involved in early development and patterning of the thyroid gland are well conserved between zebrafish and mouse (Rohr & Concha 2000, Wendl et al. 2002, Elsalini et al. 2003).
Zebrafish thyroid follicles produce thyroxine (T4) and tri-iodothyronine (T3), and two types of receptor-TRα and β have been identified (Essner et al. 1997, Liu et al. 2000), with high sequence homology to other vertebrate TRα and β receptors. As in mammals, thyroid hormone (TH) and thyroid hormone receptors (TRs) are required for correct growth and development in embryos and larval stages (Brown 1997, Power et al. 2001, Liu & Chan 2002) and also the transition from larval to juvenile stages (Brown 1997, Liu & Chan 2002) – the failure of this transition is a common problem in aquaculture of fish. In addition to its role as a model for the mammalian thyroid, the economic and conservation benefits of manipulating the thyroid function in fish are clear.
The adrenal gland
The teleost (bony fish) group to which zebrafish belong do not possess a distinct adrenal gland, but rather an intermingled group of cells sited in the anterior kidney that produce either catecholamines or steroid hormones (Gallo & Civinini 2003). The cells producing steroid hormone are referred to as the interrenals. The interrenal cells produce cortisol, which regulates both metabolism and electrolyte balance. In contrast, in terrestrial vertebrates, mineralocorticoids regulate electrolyte balance (Wendelaar Bonga 1997). As with all vertebrates, the catecholamine-producing cells are derived from embryonic neural crest cells (Reid et al. 1995), while the interrenals are derived from mesoderm (Chai et al. 2003). However, little is known about the molecular mechanisms that control inter-renal development in fish and how this compares to the mammalian adrenal gland (Chai et al. 2003).
As in other vertebrates, cortisol is produced in teleosts as a response to stress (Wendelaar Bonga 1997, Gallo & Civinini 2003), and secretion is stimulated by ACTH. In contrast to the mammalian system, cortisol secretion can also be stimulated by MSH from the pituitary (Mommsen 1999). Interrenal cells in teleosts also produce androgens (Vermeulen et al. 1994), the hormone group that can induce both oocyte maturation (Nagahama 1997) and spermiation (Todo et al. 2000) in teleosts.
The pancreas
Pancreatic development is well studied in the zebrafish model. As with mammals, it consists of both an endocrine component (the principal islet) and exocrine (parenchyma) component (Argenton et al. 1999). While molecular events appear highly conserved between zebrafish and mammals (e.g. Milewski et al. 1998, Huang et al. 2001a, Yee et al. 2001, Stafford et al. 2004, Kim et al. 2006), there are subtle differences in cellular events. The most prominent of these is that the zebrafish pancreas develops in situ from anterior endoderm cells (Ober et al. 2003, Wallace & Pack 2003), while the tetrapod pancreas forms from evagination of foregut endoderm (Pack et al. 1996).
Zebrafish genes for glucagons, insulin and somatostatin have been cloned (Milewski et al. 1998, Argenton et al. 1999, Huang et al. 2001a). There is a high level of sequence conservation with the mammalian genes, and they are expressed in pancreatic islet cells. Fish are thought to regulate blood glucose levels in the same way as mammals; that is, by glucagons and insulin (Plisetskaya & Mommsen 1996). However, there are some important differences – teleosts generally exist in a state of hyperglycaemia, even though they have insulin levels comparable to or even higher than mammals (Mommsen & Plisetskaya 1991). They have a much lower rate of glucose utilisation and a higher requirement for protein. The close link between insulin and glucagon activity in mammals may not exist in teleosts (Mommsen & Plisetskaya 1991); therefore, blood glucose may not be so tightly regulated. However, these differences may reflect the variety of environmental and metabolic differences in the aquatic environment (for review, see Moon 2001). Because of their hyperglycaemia, teleosts have been proposed as a good model for both insulin-resistant and insulin-dependent diabetes (Kelley 1993, Wilson 1994).
The gonads
Steroid hormones are synthesised in and secreted from the Leydig cells of the testis and in the ovary, under the control of pituitary gonadotrophins (FSH and LH). They are required in multiple tissues for many functions, including, in the gonad itself, regulation of the reproductive cycle. Importantly in teleosts, the developing gonad has the appearance of an immature ovary until the ‘critical period’ when sex steroids determine whether male or female organs develop (Piferrer 2001, Devlin & Nagahama 2002). In zebrafish, this influence begins at 43 dpf and is completed by 99 dpf (Maack & Segner 2004). Teleosts, including zebrafish, are used in the study of environmental endocrine disrupters due to this major influence of sex steroids on gonadogenesis. Steroidogenic synthesis of the major sex steroid appears to be conserved between mammals and fish (Fostier et al. 1983), although there are also fish-specific steroids, a fact indicating a more complex system. Of the major sex steroids, progesterone is the least studied in fish. It may stimulate gonadotrophin secretion from the pituitary at sexual maturation (Atteke et al. 2003), and a derivative, dihydroxy-4-pregnen-3-one (DHP), causes oocyte maturation in fish, including the zebrafish (Selman et al. 1994). Testosterone is the major androgen produced. Many teleosts have two androgen receptors (ARs) compared with the single mammalian AR. These teleost ARs have been shown to have high binding affinity for either testosterone or dihydrotestosterone, while the mammalian AR has high specificity for dihydrotestosterone (Sperry & Thomas 1999a, 1999b). Three oestrogen receptors (α, β1 and β2) (Bardet et al. 2002, Menuet et al. 2002) have been identified in zebrafish compared with two in mammals (α and β) (Green et al. 1986, Kuiper et al. 1996). While their sequence is homologous, zebrafish receptors differ from mammalian counterparts in response to some oestrogens (Bardet et al. 2002), a fact that, the authors suggest, may make them a less favourable model for study of endocrine disrupters. An additional site of oestrogen synthesis in all vertebrates is neural tissue (Callard 1993). Teleosts, however, have 100–1000-fold higher levels of the oestrogen-synthesising enzyme aromatase in neural tissue than mammals (Pasmanik & Callard 1985, 1988), and the activity of this enzyme is comparably high (Callard 1993, Gelinas & Callard 1993). The site of this enzyme has been located in radial glial cells in the brain (Forlano et al. 2001) that are thought to be neuroprogenitor cells. Expression studies show oestrogen receptors are expressed in the same brain regions (reviewed in Pellegrini et al. 2005). The zebrafish brain, unlike the mammalian, continues to grow during adulthood, and oestrogen may play a role in this process. Therefore, the study of oestrogen function in the brain of teleosts may have exciting possibilities for research on central nervous system neuronal repair and regeneration in mammals.
In conclusion, while developmental processes in endocrine organogenesis seem similar, particularly at the molecular level, the structure of the glands can show the greatest difference between fish and mammal. This is particularly obvious in the parathyroid/gill and adrenal/interrenal systems. Despite this, endocrine function is a well-conserved process between teleost fish and mammals, the majority of hormones, hormone receptors and activities being conserved. The main differences in function are an increase in complexity in some systems in the teleost that are of significant interest to fish biologists.
Genetic manipulation – the tools employed in zebrafish studies
While it is clear that zebrafish are a good model for mammalian/human endocrine function, the mouse model is by far the most commonly employed, as it is easily manipulated genetically. However, the zebrafish is coming to the fore as a genetic model in a wide variety of studies. Here we give an overview of the methods currently being employed in the zebrafish model, mainly in studies of development, to decipher gene function. We emphasise the distinct advantages of using these methods in this model and highlight the few examples where they have been employed to address questions of endocrine function.
Watching the defectives – fluorescent reporter lines
Significant advances have been made in many areas of genetic and biological research by the ability to generate transgenic animals. Mice are the most popular model for transgenic manipulation, as techniques to generate them are well established. However, the use of transgenic zebrafish is gaining in popularity, particularly for imaging cell behaviour during development. In recent years, there has been an explosion in the creation of fluorescent reporter zebrafish lines, where GFP or other fluorescent protein is expressed under the regulatory sequence of a specific target gene (Lin 2000). Briefly, regulatory sequence is cloned into a GFP vector and injected into the developing embryo at the one-cell stage. These fish (F0) are allowed to develop, and then mated with wild-type fish to generate an F1 population. If the promoter GFP has integrated into the germ-line, the target tissue will fluoresce, and this can be examined easily in the embryo under a fluorescent microscope. Individuals with the strongest expression are chosen to establish the line (Udvadia & Linney 2003). This allows the spatiotemporal expression patterns of these genes to be examined, in real time in the living embryo. Additionally, they can be used as a ‘marker’ for a tissue, to aid the analysis of mutant fish produced in forward and reverse gene function studies (see below and Xiao et al. 2005). Thus, lines such as insulin GFP and pdx1 GFP have been created, in which GFP is specifically expressed in the pancreas (Huang et al. 2001b), and POMC-GFP, which is expressed in the pituitary (Liu et al. 2003). These have greatly aided our understanding of both the normal development of these endocrine organs and the abnormal development in mutants. Imaging of live whole zebrafish is possible up to early larval stages (e.g. Rieger et al. 2005). The challenge is now to improve imaging after the developmental period to be able to use such lines in models of adult endocrine disease.
Making mutant fish – forward genetics
Large-scale mutagenesis screens have been employed to decipher gene function in various invertebrate models, including Caenorhabditis elegans (Hirsh & Vanderslice 1976) and Drosophila (Nusslein-Volhard & Wieschaus 1980), with impressive results. However, the ability to conduct such screens in vertebrates has been hampered by the large numbers of animals required to achieve saturation, need for laboratory space to house them and the long generation time to create recessive mutants. Zebrafish are the ideal vertebrate model in which to undertake such screens because of their relatively small size, our ability to maintain them in high density and their short generation time (3 months), allowing the analysis of several generations of mutagenised animals over a short period of time (Mullins & Nusslein-Volhard 1993). The major advantage of forward genetics is that there is no requirement for knowledge of genetic control of biological function prior to the screen, allowing the identification of novel genetic pathways. One disadvantage of note is that this type of screen reveals only unique or partially redundant functions.
ENU screens
Chemical mutagens are favoured due to their high efficiency in creating point mutations in single genes in rapidly dividing premeiotic germ cells (Singer 1983). The mutagen of choice in zebrafish is N-ethyl-N-nitrosourea (ENU), as it is the most efficient in this system (Mullins & Nusslein-Volhard 1993, Mullins et al. 1994, Solnica-Krezel et al. 1994). Typically, adult males are exposed to ENU in tank water (Mullins & Nusslein-Volhard 1993) and subsequently bred with wild-type females to generate F1 founder fish. These are then either inbred or outbred with wild-type females to generate F2 families. F2s are then mated as pairs to reveal recessive mutations in the F3 generation (Solnica-Krezel et al. 1994) (Fig. 2⇓). Once a phenotype is generated, positional cloning techniques (Talbot & Schier 1999) and candidate gene testing are employed to identify the gene that has been mutated.
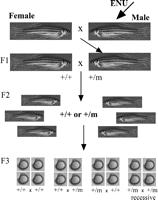
Forward genetic screens. This diagram summarises the breeding regime employed in most forward genetic screens. In the real situation, many fish are employed at each step; however, we have simplified it to make the process clearer. Male fish are mutagenised –in this case with ENU – and bred with a wild-type female to generate F1 fish, some of which carry induced point mutations. Carriers are again bred with wild-type fish to produce an increased number of carriers in the F2 family. Single pairing of sibs within the F2 families generates recessive mutants in the F3 generation, when two carriers of the mutation are mated (+/m +/m). Typically, observing embryos during development identifies recessive mutants in this F3 generation, as many have obvious aberrant morphology.
Several large-scale ENU screens have been undertaken to examine genes involved in the development of many systems (Driever et al. 1996, Haffter et al. 1996), and many smaller-scale screens have examined development in single systems (for examples, see special edition of Development, vol 123, December 1996). Some mutants of known endocrine disorders have been isolated from these screens (Table 1⇓). In addition, two screens examining the development of endocrine glands have identified 11 mutants with defects in the development of the adenohypophysis (Herzog et al. 2004b) and nine new mutants with abnormal development of pancreatic islet cells (Kim et al. 2005). The challenge now is to identify all of the mutated genes causing these phenotypes.
Human endocrine disorders phenocopied by zebrafish mutants and morphants
This type of screen has also been employed to address more diverse biochemical, physiological and pharmacological questions, such as identifying genes predisposing to cocaine addiction (Darland & Dowling 2001), and genes controlling haemostasis (Jagadeeswaran et al. 2000) and lipid metabolism (Ho et al. 2004). Of more relevance to this audience, a screen has been employed by Bauer and Goetz (2001) to examine genes involved in gonad function in sexually mature zebrafish. This has identified mutants with perturbed oocyte and sperm maturation. Thus, it is anticipated that this type of screen may be employed with significant success in endocrinology studies in the future.
Insertional screens
More recently, a large-scale, forward mutagenesis screen to identify developmental genes has been conducted by random insertion of a murine retroviral vector into the zebrafish genome (Amsterdam et al. 1999). The vector is injected into the embryo at a very early stage (a few cells), and the viral sequence is reverse transcribed and inserted into the cells’ DNA. The resulting animal is bred with a wild-type animal in a similar strategy to that described above to produce recessive mutants in the F3 generation (Fig. 2⇑). The efficiency of mutation is not as high as that with ENU, but the distinct advantage of this method is that mutated genes can be rapidly identified by RT–PCR with primers against the viral sequence (Amsterdam et al. 1999, Amsterdam 2003). Studies have also employed this method to identify genes involved in cystic kidney disease (Sun et al. 2004) and liver disease (Sadler et al. 2005).
Looking backward to go forward – reverse genetics
The large-scale sequencing of genomes, including the zebrafish, has shifted the emphasis from forward to reverse genetics. The sequence of most genes is determined and readily available, but the biological function remains to be identified (Heasman 2002). New technologies are constantly being developed to allow rapid analysis of gene function from basic sequence information. The zebrafish is at the forefront of this trend, with many simple techniques available to analyse gene function. Here we review current methods and their potential applications in endocrinology research.
Knockdown, but not knockout?
Knockouts – not yet!
One of the most successful techniques employed in understanding gene function in development and disease has been the loss-of-function knockout gene model. The establishment of stable, embryonic stem-cell lines from mice, in which targeted DNA insertion is possible, has allowed this technique to become routine in the mouse model. However, the generation of stable, heritable, targeted mutations in vertebrates other than mouse has been hampered by the absence of embryonic cell lines. The zebrafish is no exception to this, and there are currently no specifically targeted gene knockout or knockin lines. Cell lines that display ES characteristics have been generated in fish by feeder layer systems (Sun et al. 1995), but integration into the germ-cell lineage of the host embryo has been poor. However, recent advances in the derivation of cell lines that are germ-line competent over several passages and integrate into host embryos (Hong et al. 1998), and in which targeted DNA insertion appears possible and heritable (Ma et al. 2001, Fan et al. 2004), bring the prospect of targeted, knockout transgenic zebrafish a step closer to reality.
Knockdown success: morpholinos
In the absence of knockouts, knockdown strategies have come to the fore. The most widely used strategy is the application of morpholinos, antisense oligonucleotides in which the deoxyribose is substituted with an N-morpholin ring, conferring greater stability with fewer non-specific effects than other antisense DNA analogues (Summerton & Weller 1997, Corey & Abrams 2001). They bind to RNA and prevent translation, but, unlike other types of antisense oligos, do not make the RNA a substrate for RNase-H degradation. For this reason, morpholinos designed against the start site codon and 5′ UTR of a candidate gene tend to be more effective, as they stop the translational machinery (ribosome) from binding (Heasman 2002, Malicki et al. 2002). However, this is not always the case and needs to be confirmed for each gene and morpholino. They also appear to be effective if designed against splice sites (Malicki et al. 2002). They were initially designed as clinical tools (Summerton & Weller 1997) but have rapidly become a common tool in embryology, not only in zebrafish but also in other diverse models such as frog and sea urchin (reviewed in Heasman 2002).
Designing morpholinos
Morpholinos are easily designed by simple rules – they should be 25 mers, not be self-complementary, and have low guanine (G) content, with a maximum of three consecutive Gs (Malicki et al. 2002) (for further information, see www.gene-tools.com). If these rules are followed, they are readily soluble in water-based buffer such as Danieau solution (Westerfield 2000), and nanolitre volumes can be injected into the one-cell-stage embryo. Their stability ensures that their effects are long lasting. Combined with the rapid development of the zebrafish, this stability allows inhibition of translation for a large proportion of the developmental period – sometimes for 4 days (Braat et al. 2001). Phenotypes are rapidly observed in injected embryos, making this technique quick and relatively cheap to employ.
Designing morpholino experiments
There are some important considerations in performing and interpreting data from morpholino studies. A dose–response experiment is required to determine the dose at which a specific phenotype occurs and that at which toxicity occurs (as manifest, for example, by widespread cell death). It is possible that the difference between these two doses is very small. Injected morpholinos are unlikely to exhibit complete loss of function, more typically generating a graded severity of phenotype. This may be seen as a distinct advantage where complete null results in early lethality. Similarly, some are highly efficient, producing phenotype in over 90% of injected embryos, while others produce phenotype in less than 20% (reviewed in Heasman 2002).
As with all antisense experiments, effective controls must be employed. Typically, two adjacent but non-overlapping morpholinos are used to confirm that the phenotype is gene specific (Sumanas et al. 2001). Five base pair mismatched controls are also used to determine whether the phenotype produced is non-specific. The best assay to confirm the activity and specificity of the morpholino is Western blotting or immunohistochemistry – both of which require a specific primary antibody. As mRNA is not degraded, RT–PCR cannot be used as an assay in standard morpholino experiments, although it will detect smaller transcripts if the morpholino is directed against a splice site (Malicki et al. 2002). In the absence of an antibody, specificity can be demonstrated by rescue of the phenotype by injection of synthetic mRNA. However, this must either lack the 5′UTR sequence or have minor base-pair modification to stop morpholino binding and inactivation of the injected mRNA (Nasevicius & Ekker 2000, Cui et al. 2001).
Application of morpholinos in endocrine studies
There is now a wealth of published evidence to show the effectiveness and specificity of morpholinos in zebrafish. Their use in endocrine research is likely to predominate in developmental studies, and candidate genes that are proposed to play a role in endocrine gland development have already been examined by this system (Table 1⇑). For example, morpholinos against Pdx-1 were employed to demonstrate a role for this gene in endocrine pancreas formation (Huang et al. 2001a, Yee et al. 2001). Similarly, in the developing interrenal cells (adrenal gland homologue), morpholinos against ff1b (the homologue of mammalian SF-1 gene) disrupted interrenal development, downregulated expression of genes coding for steroidogenic enzymes, and impaired osmoregulation in the larval fish (Chai et al. 2003), partially phenocopying human and mouse mutations in the same gene. Injection of morpholinos targeted against the gene Pit1 resulted in loss of the lactotroph, somatotroph and thyrotrophic cells in the developing pituitary and a lack of growth in the juvenile (dwarfism) (Nica et al. 2004). Importantly, this morpholino mutant phenocopies zebrafish Pit1 mutants isolated in an ENU screen (Herzog et al. 2004b, Nica et al. 2004), mouse Pit1 mutants (Snell dwarf mouse (Li et al. 1990)) and human Pit1 mutants (combined pituitary hormone deficiencies (CPHD) (Pfaffle et al. 1992, Radovick et al. 1992, Tatsumi et al. 1992), indicating the potential for such studies in examining endocrine development and function. To contrast with this, morpholinos have also been used to demonstrate a role for the endocrine hormones GnRH, GHRH and PACAP during development (Sherwood & Wu 2005).
These studies demonstrate the validity of the morpholino system, and in view also of their cost-effectiveness and ease of use, morpholinos will be a valuable tool for deciphering both the genetic control of endocrine gland development and the developmental effects of endocrine hormones in the future.
Gaining ground – gain of function studies
Overexpression of genes is another way of defining gene function. As with knockdown studies, a candidate gene must be identified. However, it can have distinct advantages over knockdown, including the ability to express protein subunits or mutated proteins to decipher structure–function relationships. Overexpression of genes in zebrafish embryos is easily and routinely performed and often performed in parallel with knockdown studies to decipher gene function.
The most common method is by injection of capped synthetic mRNA into the developing embryo. mRNA is transcribed in vitro from a DNA construct and capped to increase efficiently of translation in the developing embryo. Injection of mRNA at a concentration of 10–100 μg/ml is sufficient to cause uniform expression (Malicki et al. 2002). For example, this technique has been used to assign a role for thyroid hormone receptor alpha 1 in early neural development (Essner et al. 1999) and to identify a role for the Sonic hedgehog (shh) gene in adenohypophysis development (Herzog et al. 2003).
Temporal control of overexpression is often desirable, as most developmentally important genes have multiple functions at different times of development (Malicki et al. 2002). This is now being made possible by the availability of heat-shock-inducible vectors that can be injected into the embryo; however, a potential problem may lie in the fact that such vectors show a mosaic distribution in the embryo, and thus ubiquitous and uniform overexpression may be difficult to achieve (Riley et al. 2004). Currently, inducible gene expression systems such as this are not widely used in the adult animal, and that is a distinct disadvantage to many endocrinologists.
TILLING your way to the top
Both forward and reverse genetic screens have contributed significantly to our understanding of gene function in development and disease. However, they both have distinct disadvantages. Mutational screens are efficient and require no prior information, but cloning the gene mutated can be laborious, and the mutant line has to be maintained during this investigation, or the sperm must be frozen. Reverse genetic techniques, such as morpholinos, are quick and simple but require a candidate, and are not germ-line mutations, nor are they stable over protracted periods of time. This, coupled with the inability to create targeted knockouts, may make zebrafish seem a less attractive genetic system. However, the combination of forward and reverse genetics appears to have overcome these disadvantages to generate knockout lines for the research community by the TILLING (targeted, induced local lesions in genomes) technique (McCallum et al. 2000). This combines a classical high-saturation ENU screen with high-throughput screening for point mutations in the genome. It was initially developed as a technique in plant genetics (Henikoff et al. 2004) but has been applied to animal models including zebrafish. After ENU mutagenesis, genomic DNA is harvested and rapidly screened, using RT–PCR and a range of primers designed against known target genes. This strategy can detect single mutations in DNA, some of which will be null mutations (Wienholds et al. 2002). Instead of sequencing all samples to detect mutations, PCR products are denatured and reannealed to form heteroduplexes, which are then cleaved by the enzyme CEL-1. Samples are analysed on a denaturing polyacrylamide gel, and potential null mutants are selected and sequenced (Wienholds et al. 2003). This method is rapid, allows screening of many samples at a time (Stemple 2004) and results in the production of a founder fish for a ‘knockout’ line for a given candidate gene. As an added benefit, this method allows the production of ‘living libraries’ where the useful mutation is identified while the mutagenised fish are still reproductively viable, and stops the loss of alleles associated with cryopreservation (Wienholds et al. 2003). Several zebrafish TILLING projects are under way, and it is anticipated that they will generate a variety of mutants of interest to the wider endocrinology community.
Techniques for the future – transposons and gene traps
Recent papers have discussed the use of transposons in zebrafish genetics. Transposons can be inserted into the germ line easily and have been used to great effect in the enhancer gene trap method in Drosophila (O’Kane & Gehring 1987) and mouse (Friedrich & Soriano 1991). Zebrafish are the ideal vertebrate for enhancer gene traps, as they can be easily analysed for expression patterns through simple fluorescent microscopy in the embryo. Two groups have recently used transposons as a gene trap to create lines with GFP expression in specific tissues far quicker than the current transgenic method (Balciunas et al. 2004, Kawakami et al. 2004). The exciting prospect is the development of this system into a method to create insertional mutants similar to those of the UAS-Gal4 system in Drosophila, which has proved so successful in determining gene function in that model.
Conclusion
It is clear that the zebrafish is a good model for vertebrate endocrine function. In addition, it should not be ignored that the study of fish endocrinology is of important scientific, economic and ecological benefit. The advances in genome sequencing and the techniques available to manipulate gene function make the zebrafish an attractive model for scientific research. Some areas of endocrine research, most notably endocrine gland development, have begun to take advantage of this model. Currently, the biggest disadvantage of the zebrafish in endocrine research is the lack of cell lines. However, as the use of this model increases, it is anticipated that these will be developed. Similarly, with the advent of more sophisticated techniques to control spatiotemporal loss or gain of function, such as caged morpholinos and inducible gene expression systems that work in both embryo and adult, it is hoped that this use will rapidly spread into other areas of endocrine research and that zebrafish will become the model of choice in the coming years.
- Received 2 December 2005
- Accepted 20 January 2006
- Accepted 30 January 2006
- Made available online as an Accepted Preprint 30 January 2006
- © 2006 Society for Endocrinology










