Developmental regulation of the lung in preparation for life after birth: hormonal and nutritional manipulation of local glucocorticoid action and uncoupling protein–2
- Centre for Reproduction and Early Life, Institute of Clinical Research, University of Nottingham, Nottingham NG7 2UH, UK
- (Requests for offprints should be addressed to M E Symonds; Academic Division of Child Health, School of Human Development, Queen’s Medical Centre, University Hospital, Nottingham NG7 2UH, UK; Email: michael.symonds{at}nottingham.ac.uk)
Abstract
Glucocorticoid action has a major role in regulating fetal and postnatal lung development, although its impact on mitochondrial development is less well understood. Critically, the consequences of any change in glucocorticoid action and mitochondrial function in early life may not be limited to the postnatal period, but may extend into later life. This paper focuses on more recent findings on the impact of ontogeny, fetal cortisol status, maternal nutrient restriction and postnatal leptin administration on mitochondrial uncoupling protein (UCP)-2, glucocorticoid receptor (GR) and 11 β-hydroxysteroid dehydrogenase type 1 (11βHSD1) isoform abundance in the lung. For example, in sheep, GR and 11βHSD1 mRNA are maximal at 140 days’ gestation (term ~147 days), while UCP2 mRNA peaks at 1 day after birth and then decreases with advancing age. In the fetus, chronic umbilical cord compression enhances the abundance of these genes, an outcome that can also be produced after birth following chronic, but not acute, leptin administration. Irrespective of the timing of maternal nutrient restriction in pregnancy, glucocorticoid sensitivity and UCP2 abundance are both upregulated in the lungs of the resulting offspring. In conclusion, prenatal and postnatal endocrine challenges have distinct effects on mitochondrial development in the lung resulting from changes in glucocorticoid action, which can persist into later life. As a consequence, changes in glucocorticoid sensitivity and mitochondrial protein abundance have the potential to be used to identify those at greatest risk of developing later lung disease.
Introduction
Mitochondria, the ‘powerhouses’ of the cell, control the oxidative energy supply in normal and pathological physiology, embryonic development, apoptosis and general body ageing (Gottlieb 2000). While the metabolic regulation of mitochondria is well described, the endocrine and nutritional regulation of specific mitochondrial proteins, particularly during fetal and postnatal development, is less well defined. This is perhaps surprising given the very high metabolic demands on the newborn that have a considerable impact on both lung and mitochondrial function (Symonds et al. 1993, 1995, 2003).
The transition from fetus to neonate represents a major physiological, hormonal and environmental challenge. In order to survive, the fetus must establish independent thermoregulation and ventilation shortly after birth, and this requires a number of highly complex physical and hormonal interactions around the time of parturition. Effective ventilation after birth is dependent on the rapid activation of lung function, which is controlled by a number of hormones during the peripartum period (Grier & Halliday 2004, Rajatapiti et al. 2005). These help to ensure that there is rapid clearance of lung fluid followed by the entry of air processes, which can occur independently (Olver et al. 2004). Our understanding of the endocrine and nutritional regulation of lung development mainly stems from studies utilising the sheep, undoubtedly the species of choice to investigate these variables. The newborn sheep, like the human infant, has a mature hypothalamic-pituitary-thyroid-adrenal axis at birth, while there is a marked difference in lung development and mitochondrial abundance between rodents and larger mammals (Gnanalingham et al. 2005a). In sheep, total mitochondrial protein abundance increases during gestation to peak around the time of birth in the majority of tissues, including the lung, studied to date (Mostyn et al. 2003b).
In the following review, we will focus on the effect of age (ontogeny); discrete nutritional and endocrine modulation of specific lung mitochondrial proteins and receptors, namely, uncoupling protein (UCP)-2, voltage-dependent anion channel (VDAC), cytochrome c; and local glucocorticoid action, as determined by the abundance of glucocorticoid receptor (GR) and isoforms of 11 β-hydroxysteroid dehydrogenase (11βHSD). The locations and postulated roles of UCP2, VDAC and cytochrome c are outlined in Fig. 1⇓.
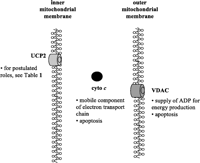
Schematic diagram of mitochondrial protein location and function in the lung. UCP: uncoupling protein; VDAC: voltage-dependent anion channel; cyto c: cytochrome c; ADP: adenosine diphosphate. Adapted from Mostyn et al.(2004).
Mitochondrial proteins and energy metabolism
Uncoupling proteins are members of the inner mitochondrial membrane carrier subfamily. UCP2 shares 56% homology with UCP1 (Erlanson-Albertsson 2003), and is widely expressed in white adipose tissue, lung, spleen, thymus, macrophages, bone marrow, stomach, islets of Langerhans, skeletal muscle, heart, kidney, placenta and brain (Ricquier & Bouillaud 2000, Pecqueur et al. 2001). This recognised wide tissue distribution is based primarily on UCP2 mRNA determination, while the localisation of the protein has proved more problematic, mainly because of the lack of antibodies sufficiently sensitive to detect the protein, which, in adults, is present at 1% of the levels of UCP1 in brown adipose tissue (Erlanson-Albertsson 2003). Using specific antibodies raised against the human sequence, UCP2 protein was primarily located in white adipose tissue, lung, spleen and stomach (Pecqueur et al. 2001). The marked discrepancy in UCP2 mRNA and protein expression potentially reflects the translational regulation of UCP2 mRNA by an upstream open reading frame located in exon 2 of the UCP2 gene, which strongly inhibits the expression of the protein (Pecqueur et al. 2001). Consequently, the exact function of UCP2 remains a subject of intense debate (Nedergaard & Cannon 2003), with postulated roles in energy balance (Buemann et al. 2001), reactive oxygen species (ROS) production (Negre-Salvayre et al. 1997) and apoptosis (Voehringer et al. 2000), as summarised in Table 1⇓.
Postulated roles for uncoupling protein-2
Located in the outer mitochondrial membrane, VDAC, an anion selective channel, is considered to be the main pathway for metabolite and ion diffusion across this membrane due to its propensity to form pores (Colombini 1979). Respiration in mitochondria may be influenced by changes in the permeability of the outer membrane regulated by VDAC (McEnery et al. 1993), although it has a range of functions dependent on tissue location and may also be present in other extramitochondrial membranes (Shinohara et al. 2000). VDAC, along with other membrane proteins, could also be responsible for the release of cytochrome c from the intermembrane space, a process that has been implicated in the chain of events involved in apoptosis (Crompton 1999). Cytochrome c is itself an essential component of the mitochondrial respiratory chain and is a mobile electron transporter, involved in the electron transfer from complexes III and IV of the electron transport chain (Lehninger et al. 1993).
Local glucocorticoid action
Tissue sensitivity to glucocorticoids is regulated predominantly by intracellular expression of the GR and 11β-hydroxysteroid dehydrogenase type 1 (11βHSD1) and type 2 (11βHSD2) at the level of gene transcription. The GR is found in almost every cell in the body and serves a multitude of physiological functions, ranging from glycogen deposition in the liver to inhibition of lymphocyte function (Svec & Rudis 1981, Sapolsky et al. 1984). Glucocorticoids can modulate their own action, not only by inhibition of the hypothalamic-pituitary axis, but also by a negative feedback downregulation of GR number (Svec & Rudis 1981, Sapolsky et al. 1984). Changes in GR response can also be important, either at the central nervous system level, by modulating the negative glucocorticoid feedback (De Kloet et al. 1998), or at a peripheral level, by regulating cell differentiation and metabolism in a tissue-specific fashion (Kissebah & Krakower 1994). The critical importance of the GR in survival after birth is illustrated by mice that lack the GR gene, most of which (> 90%) die at birth from respiratory dysfunction (Cole et al. 1995). Not surprisingly, lung development in terms of its gross structure, as well as epithelial cell structure, is severely retarded in these fetuses (Cole et al. 2004).
In the developing lung, 11βHSD1 and 2 closely follow changes in the GC (Gnanalingham et al. 2005a). 11βHSD1 behaves predominantly as an 11-oxoreductase, utilising nicotinamide adenine dinucleotide (NAD) phosphate as a cofactor to catalyse the conversion of inactive cortisone to bioactive cortisol, and as an intracellular modulator of glucocorticoid excess to the GR (Bamberger et al. 1996, Stewart & Krozowski 1999). It is widely distributed and present in lung as well as adipose tissue, skeletal muscle, heart, brain and adrenal cortex (Seckl et al. 2004). Conversely, 11βHSD2 behaves as an NAD-dependent dehydrogenase, catalysing the inactivation of cortisol to cortisone, and thereby maintains the specificity of the mineralocorticoid receptor for aldo-sterone (Stewart & Krozowski 1999). 11βHSD2 is generally restricted to mineralocorticoid target tissues, such as the kidney, sweat glands, salivary glands and gastric mucosa (Seckl et al. 2004), but is also highly abundant in the lung (Gnanalingham et al. 2005a).
Changes in the lung during fetal and postnatal life
In man and sheep, the development of lung architecture occurs during fetal and early postnatal life, in four defined phases (embryonic, pseudoglandular, canalicular and alveolar; Fig. 2⇓), which depend partly on intact fetal adrenal and thyroid glands and the associated prepartum surge in fetal plasma cortisol and tri-iodothyronine (T3) (Symonds & Clarke 1996, Fowden et al. 1998). In addition, fetal plasma catecholamines, with noradrenaline constituting 40–60% of total catecholamines in the fetal sheep adrenal gland (Robinson et al. 1983), are needed for increased surfactant production and decreased lung fluid production near term (Phillippe 1983, Wallace et al. 1996). By the time of birth, the airways have been laid down and the formation of alveoli has begun, although it is clear that, at birth, gas exchange may largely take place within terminal air sacs, the predecessors of definitive alveoli. After birth, more alveoli are formed until the age of 18–36 months in offspring (Thulbeck 1982), after which lung growth occurs largely by enlargement of existing alveoli. Thus, it is evident that environmental compromises imposed during fetal or early postnatal life, when lung architecture is being laid down, can result in permanent alterations in lung structure that have the potential to affect later function (Harding 1994).
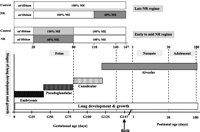
Phases of fetal lung development and postnatal lung growth in relation to the timing of periods of maternal nutrient restriction in the sheep. The phases of fetal lung development are as follows: embryonic, 0–40 days’ gestation; pseudoglandular, 40–80 days; canalicular, 80–120 days; alveolar, 120 days to term (148 days). In early to middle (28–80 days’ gestation) maternal nutrient restriction (NR), lungs were sampled in the fetus at 80- and 140-day gestation and sheep at 180 days (6 months), while, in late (110–147 days’ gestation) maternal NR, lungs were sampled at 1 and 30 days’ postnatal age. NR mothers received 60% of their metabolisable energy (ME) requirements for maternal metabolism and fetal growth; control mothers received 100% (late gestational NR) to 150% (early to midgestational NR) ME requirements during the period of NR. G: gestational age. Adapted from Alcorn et al.(1981), Harding (1994) and Gnanalingham et al. (2005b).
In the fetus, lung growth can be affected by metabolic, endocrine and inflammatory factors affecting the intrauterine environment (Kotecha 2000). These are not mutually exclusive, in that alterations in one may affect others. Indeed, several are associated with intrauterine conditions that lead to low birth weight, by restricting fetal growth and/or leading to preterm birth, including fetal nutrient restriction, fetal hypoxaemia, elevated circulating corticosteroid levels and intrauterine infection (Harding et al. 2000). They also affect mitochondrial protein abundance in the lung (Gnanalingham et al. 2005a, 2005b, Mostyn et al. 2003b).
In the sheep lung, the peak in UCP2 mRNA at day 1 precedes that of the protein at day 7 of age, coinciding with maximal abundance of VDAC, while cytochrome c protein abundance remains unchanged with age (Mostyn et al. 2003b, Gnanalingham et al. 2005a). The developmental ontogeny of UCP2 mRNA in the sheep lung appears to be markedly different from that in the rodent, where the concentration of UCP2 mRNA is low and unchanged during late gestation, doubling within 6 h after birth and remaining high to adulthood (Xiao et al. 2004). Furthermore, in contrast to rodents (Pecqueur et al. 2001), UCP2 mRNA correlates with protein levels in the postnatal sheep lung (Mostyn et al. 2003b, Gnanalingham et al. 2005a). These changes in UCP2 closely follow the peak abundance in GR and 11βHSD1 mRNA prior to term, thereby indicating a potential developmental link between UCP2 and cortisol (Gnanalingham et al. 2005a) (Fig. 3⇓). Taken together, such findings suggest a potential role for UCP2 and local glucocorticoid action within the lung in the peripartum period, mirroring the developmental ontogeny of brown adipose tissue-specific UCP1, which is primarily involved in non-shivering thermogenesis (Clarke et al. 1997b). A direct thermogenic role for UCP2, however, appears very unlikely, considering the low guanosine diphosphate (GDP)-binding activity in the fetal and postnatal lung, which is approximately 5% of that found in newborn brown adipose tissue (Gnanalingham et al. 2005a). Potential glucocorticoid and thyroid response elements have been identified in the promoter region of human UCP2 (Tu et al. 1999), suggesting that the developmental ontogeny of UCP2 mRNA expression in the sheep lung could be directly or indirectly regulated by glucocorticoids and thyroid hormones, as is the case for UCP1 (Mostyn et al. 2003a). Indeed, a range of mitochondrial proteins and cellular enzymes in the lung are influenced by the fetal endocrine environment (Mostyn et al. 2003b, Gnanalingham et al. 2005a, 2005b).
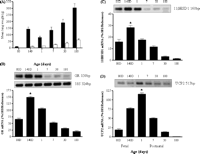
Ontogeny of (A) fresh ▪ and dry lung weights □, (B) glucocorticoid receptor (GR) mRNA, (C) 11β-hydroxysteroid dehydrogenase type 1 (11βHSD1) mRNA and (D) uncoupling protein-2 (UCP2) mRNA, between the fetus at 80- and 140-day gestation (term ~147 days) and 6 months’ postnatal age in the sheep lung. Examples of each gene mRNA expression are given. 18S, 18S mRNA. Values are means ± s.e. (n = 6 per time point). *Maximal abundance detected, significantly different (P < 0.01) from all other age groups, and greatest at 180 days (6 months). Adapted with permission from Gnanalingham et al. (2005a).
Endocrine manipulation of fetal lung mitochondrial development
During late gestation, a number of metabolic hormones have significant effects on fetal lung mitochondrial maturation, although cortisol appears to be the most important. For example, fetal plasma cortisol is raised after umbilical cord compression during late gestation (Gnanalingham et al. 2005b). In man, umbilical cord compression occurs in up to 40% of pregnancies and can result from nuchal cord (Capeless & Clapp 1989), torsion of the umbilicus during gestation (Rayburn et al. 1981), oligohydramnios (Leveno et al. 1984) or compression of the cord during the actual processes of labour and delivery (Wheeler & Greene 1975). It increases the susceptibility of the fetus to perinatal complications and, potentially, neurodevelopmental handicap (Mann 1986, Capeless & Clapp 1989). In fetal sheep, partial compression of the umbilical cord to reduce umbilical blood flow by 30% from baseline for a period of 3 days produces reversible mild fetal asphyxia, a transient increase in fetal plasma adrenocorticotrophic hormone (ACTH) and catecholamines, and a progressive and sustained increase in plasma cortisol, without affecting fetal plasma thyroid hormone concentrations (Gardner et al. 2001, Gnanalingham et al. 2005b). The low fetal arterial pH and oxygen partial pressure, and increased fetal arterial carbon dioxide partial pressure evident during umbilical cord compression, may affect and/or oppose the ability of cortisol to stimulate thyroid hormone bioavailability (Thomas et al. 1978, Wu et al. 1978).
Chronic umbilical cord compression results in upregulation of UCP2 mRNA, and VDAC and cytochrome c protein abundance with differential effects on local glucocorticoid action and GDP binding (Gnanalingham et al. 2005b). It causes an increase in 11βHSD1 mRNA, but decreased 11βHSD2 mRNA abundance and GDP binding in the lung. Furthermore, UCP2, GR and 11 βHSD types 1 and 2 mRNA, as well as VDAC and cytochrome c protein abundance, are all significantly correlated with fetal plasma cortisol and catecholamine levels, but not thyroid hormone concentrations, in the fetal lung of umbilical cord compression fetuses (Gnanalingham et al. 2005b). These parallel changes in the mitochondria within fetal lung may better prepare the compromised fetus for preterm birth and extrauterine adaptation by establishing and maintaining effective ventilation. To this extent, VDAC is located within the lung bronchioles of the fetal lung (Yakubu 2005), whereas the exact location of UCP2 remains to be confirmed due to the current unavailability of specific antibodies for ovine UCP2 (Gnanalingham et al. 2005c).
Effects of leptin administration on neonatal lung mitochondrial development
While it is now established that the fetal endocrine environment affects mitochondrial development in fetal lung, our knowledge of the endocrine milieu postnatally is still limited, despite the fact that this environment affects the extrauterine adaptation necessary to maintain effective ventilation in the neonate (Clarke et al. 1997c). A potential endocrine factor that may be important in lung development is leptin, a 16 kDa polypeptide hormone, principally synthesised and secreted by adipose tissue, which acts to regulate energy homeostasis and a range of neuroendocrine and reproductive functions in the adult (Friedman & Halaas 1998, Ahima & Flier 2000). Although the exact role of leptin in the neonate has not been fully established, leptin has been recently implicated in newborn thermogenesis in large mammals (Mostyn et al. 2002, 2004) and in lung growth (Tsuchiya et al. 1999).
Cumulative evidence has identified both fetal and adult lungs as leptin-responsive (Tsuchiya et al. 1999, Henson et al. 2004), and shown that leptin regulates immune function (Loffreda et al. 1998, Lord et al. 1998). UCP2-deficient mice are resistant to infection with Toxoplasma gondii, and their macrophages generate 80% more ROS than wild-type mice and in vitro have fivefold greater toxoplasmacidal activity, which is absent in the presence of a quencher of ROS (Arsenijevic et al. 2000). This proposed role for UCP2 in macrophage-mediated immunity and limitation of ROS has been supported by others (Negre-Salvayre et al. 1997, Ookawara et al. 2002), although this role has not been confirmed in vitro with mitochondria isolated from the lung of UCP2 knockout mice (Couplan et al. 2002).
Decreased abundance of UCP2 plus increased glucocorticoid action in the lung with chronic, but not acute, leptin administration (Gnanalingham et al. 2005c) could promote ROS production and maintain host immunity through augmentation of alveolar macrophage phagocytosis and leukotriene synthesis (Arsenijevic et al. 2000, Mancuso et al. 2002). The impact of a precocious decrease in UCP2 with leptin administration on later lung function has yet to be examined, but it may be important in tissue development, thereby optimising lung function in the neonate. Indeed, it may increase the epithelial response to both inspired air as well as potentially harmful pathogens, although this remains to be examined.
Maternal nutrition and fetal programming of the lung
At least 30% of annual deaths in the world are result from cardiovascular disease (CVD), including hypertension, type II diabetes and obesity, as well as lung disease. Extensive epidemiological evidence indicates that the nutritional and hormonal environment encountered by the fetus is a strong determinant of not only fetal growth but also disease risk in later life (Barker 1998, 1999). As a result of these adaptations, infants born to mothers that are nutrient restricted in utero are biologically different from infants born to adequately nourished mothers (Barker et al. 2001). These epidemiological findings are now strongly supported by experimental animal studies indicating that fetal physiological development and organ sensitivity, including that of the lung, are significantly altered by maternal nutrient restriction (Symonds et al. 1995). Diet type and composition, frequency of feeding, maternal age and maturity of offspring may further influence these adaptations at birth. Such adaptations have the potential to contribute to a range of diseases, including chronic lung disease, in later life.
Maternal nutrient restriction between early and midgestation
The Dutch famine of 1944–5 led to one of the most striking epidemiological findings relating to the timing of maternal nutrient restriction and subsequent increase risk of obstructive airways disease in adulthood (Lopuhaa et al. 2000), although those victims that reported a greater incidence of lung disease showed no measurable change in measured lung function. During the 5-month period of the famine, mean energy intake was 3.2 MJ/day compared with 6.3 MJ/day immediately afterward (Roseboom 2000). Dietary restriction during early gestation was shown to have the greatest effect on placental size:birth weight ratio, and to result in a much greater risk of adult CVD, obesity and obstructive airways disease (Lopuhaa et al. 2000, Roseboom et al. 2000a, 2000b), but, to date, the victims’ overall rate of mortality has remained unaffected, at least up to 57 years of age (Painter et al. 2005). Interestingly, sheep models of undernutrition that broadly adopt the same magnitude of maternal nutrient restriction as that imposed under the Dutch famine seldom affect birth or tissue weight, but have a pronounced impact on placental mass (Dandrea et al. 2001). The consistent finding that 50% variation in maternal food intake can determine placental mass may have further relevance to contemporary human populations, for which a similar range between the upper and lower quartiles in energy intake is found in both early and late gestation (Godfrey et al. 1996).
In the sheep lung, maternal nutrient restriction in early to midgestation increased UCP2 mRNA and local glucocorticoid action in the fetus, adaptations that were still evident at 6 months of age (Whorwood et al. 2001, Gnanalingham et al. 2005a) (Fig. 4⇓). Although the exact mechanisms by which maternal nutrient restriction upregulates UCP2 in the lung have yet to be determined, both non-esterified fatty acids (NEFA) and glutathione have postulated roles. In both adult and neonatal rodents, NEFA have been proposed to regulate lung UCP2 mRNA, since calorie restriction causes a rapid increase in NEFA, and lung UCP2 mRNA is increased by NEFA administration to fed animals (Xiao et al. 2004). In sheep, while there is an increase in maternal plasma NEFA between 80 and 140 days’ gestation, there are no differences in maternal or fetal plasma NEFA after 80 days’ gestation following maternal nutrient restriction in early to midgestation (Bispham et al. 2003), and maternal NEFA do not cross the ovine placenta.
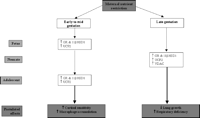
Summary of the effects of early to mid- and late-maternal nutrient restriction on mitochondrial proteins and cellular enzymes in the lung. UCP2: uncoupling protein-2; GR: glucocorticoid receptor; 11βHSD: 11β-hydroxysteroid dehydrogenase; VDAC: voltage-dependent anion channel.
Another potential nutritional mediator is glutathione, for which there is a marked reduction in plasma concentrations with starvation (Smith & Anderson 1992). This may upregulate UCP2 expression in the lung by increasing levels of intracellular ROS (Pecqueur et al. 2001). A similar mechanism has been proposed for the increase in lung UCP2 with lipopolysaccharide injection, whereby macrophage receptors stimulate the production of proinflammatory cytokines, such as tumour necrosis factor (TNF)-α (Clarke et al. 1997a), which activates the nuclear factor-κB pathway, again increasing levels of intracellular ROS (Pecqueur et al. 2001). It is uncertain, however, what effect glutathione has on the fetus at the time of nutrient restriction, particularly within the lung.
The increase in UCP2 mRNA and local glucocorticoid action that follows maternal nutrient restriction may potentially underlie some of the detrimental effects on fetal and postnatal lung development, although this remains to be fully established. Previous studies of developing animals have shown that both prenatal and early postnatal undernutrition can result in alterations in lung structure and function, with additional adverse effects on respiratory muscle development (Prakash et al. 1993, Sahebjami 1993). After restriction of placental growth, the tracheas of growth-restricted sheep fetuses were found to have thinner mucosal and submucosal layers, reduced ciliation of epithelial cells, and reduced extent of mucosal folding near term (Rees et al. 1991). Children with evidence of growth restriction in utero have reduced forced expiratory flow rates, indicative of impaired airway function (Nikolajev et al. 1998, Magness & Poston 2005), a condition persisting into adulthood with increased risk of respiratory morbidity and mortality (Barker et al. 1991, Stein et al. 1997). While the molecular bases for these detrimental effects on lung development and function have to be fully elucidated, there is evidence that a cascade of inflammatory markers, including TNF-α and interleukin (IL)-1β, which are also known to upregulate UCP2 in the lung (Pecqueur et al. 2001), may underlie the development of chronic lung disease (Ozdemir et al. 1997, Allen 2003). The increased UCP2 mRNA abundance within the lung after maternal nutrient restriction in early to midgestation may predispose to the development of chronic lung disease, especially since UCP2 plays a role in intracellular ROS production (Negre-Salvayre et al. 1997), apoptosis (Voehringer et al. 2000) and macrophage-mediated immunity (Arsenijevic et al. 2000).
Maternal nutrient restriction in late gestation
Compensatory changes in mitochondrial protein and receptor abundance within the lung after maternal nutrient restriction during late gestation are evident. VDAC abundance in nutrient-restricted offspring is enhanced up to at least 1 month of age (Mostyn et al. 2003b). There is an accompanying increase in UCP2, GR and 11βHSD1 mRNA abundance up to 1 month of age after maternal nutrient restriction in late gestation (Gnanalingham et al. 2005a) (Fig. 4⇑). Interestingly, adaptations of this type in adipose tissue can affect obesity (Seckl et al. 2004); in sheep, they follow the increase in fat mass with age (Bispham et al. 2005). Also in sheep, maternal nutrient restriction in late gestation reduces lung growth (Harding & Johnston 1995, Symonds et al. 1995) and impairs the growth of the bronchial walls, possibly affecting airway compliance in the immediate postnatal period (Wignarajah et al. 2002). Increased UCP2 mRNA abundance within the lung may underlie the impairment of lung growth and function, by increased intracellular ROS production, activation of macrophages and secretion of proinflammatory cytokines, including TNF-α and IL-1β (Pecqueur et al. 2001, Alves-Guerra et al. 2003). Impaired pulmonary defence mechanisms may result from undernutrition (Bellanti et al. 1997), possibly contributing to greater susceptibility to respiratory infections, especially since the development of mucous elements remains altered in the postnatal lung after restricted fetal growth in late gestation (Wignarajah et al. 2002). The enhanced UCP2 mRNA abundance may also increase susceptibility to infection and death, as demonstrated with Toxoplasma gondii (Arsenijevic et al. 2000).
The mechanism by which maternal nutrient restriction during late gestation affects lung mitochondrial development has yet to be fully determined. Variations in fetal plasma cortisol and thyroid hormones may not be directly responsible for these tissue-specific responses after maternal undernutrition during late gestation. In sheep, maternal cortisol is increased for only the first 10 days of nutrient restriction when food intake is reduced from 110 days’ gestation and there is no change in fetal plasma cortisol concentration (Edwards & McMillen 2001). Although the increase in maternal and fetal plasma thyroxine with gestational age toward term (Fraser & Liggins 1989, Bispham et al. 2003) may contribute to the ontogenic changes in local glucocorticoid action and UCP2 mRNA abundance within the lung during the peripartum period, both maternal and fetal plasma thyroid hormone concentrations are reduced by maternal nutrient restriction (Rae et al. 2002, Bispham et al. 2003). Hence, the observed changes in local glucocorticoid action and UCP2 mRNA abundance in the lung occur in the absence of any consistent changes in maternal or fetal plasma hormone concentrations after late maternal nutrient restriction.
Conclusion
Pre- and postnatal challenges have distinct effects on lung mitochondrial development (Fig. 5⇓), which can persist into later life. A greater understanding of the regulation of these mitochondrial proteins and local glucocorticoid action within the lung by endocrine and nutritional factors is likely to be important in preventing neonatal morbidity and mortality. The potential ‘programming’ of these tissues at the molecular level may underlie the physiological and pathophysiological processes implicated in disease development, including chronic lung disease. Changes in these mitochondrial proteins may therefore provide a potential marker for identifying those most at risk of developing adulthood disease and potential targets of therapeutics.
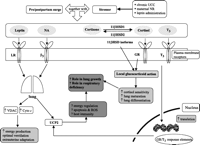
Summary of the main regulators of local glucocorticoid action and UCP2 within the lung. UCC: umbilical cord compression; NR: nutrient restriction; ROS: reactive oxygen species; LR: leptin receptor; GR: glucocorticoid receptor; β3: β3-adrenergic receptor; NA: noradrenaline; T3: tri-iodothyronine; UCP2: uncoupling protein-2; VDAC: voltage-dependent anion channel; cyto c: cytochrome c; 11βHSD1: 11β-hydroxysteroid dehydrogenase type 1; 11βHSD2: 11β-hydroxysteroid dehydrogenase type 2.
Acknowledgments
We thank the Special Trustees for Nottingham University Hospitals and the British Heart Foundation for financial support of this work. The authors declare that there is no conflict of interest that would prejudice the impartiality of this scientific work.
- Received in final form 15 December 2005
- Accepted 5 January 2006
- Accepted 19 January 2006
- Made available online as an Accepted Preprint 19 January 2006
- © 2006 Society for Endocrinology










