| HOME | HELP | CONTACT US | SUBSCRIPTIONS | ARCHIVE | SEARCH | TABLE OF CONTENTS |
|
|
||||||||
STARLING REVIEW |  |
Institute of Reproductive and Developmental Biology, Faculty of Medicine, Imperial College, Du Cane Road, London W12 0NN, UK
(Requests for offprints should be addressed to J H Steel; Email: j.steel{at}imperial.ac.uk)
| Abstract |
|---|
|
|
|---|
| Introduction |
|---|
|
|
|---|
RIP140 binds to nuclear receptors in a ligand-dependent manner by a mechanism which resembles that of the p160 coactivators. These cofactors contain helical leucine-rich LXXLL motifs that dock into a cleft formed by the activation surface in the ligand-binding domain of nuclear receptors when a hormonal ligand is bound (Heery et al. 1997, Torchia et al. 1997). The mechanisms that determine whether, for example, a p160 coactivator or the RIP140 corepressor is recruited to a particular nuclear receptor and precisely when and for how long are not known but there are hints that several mechanisms may be involved. These include variations in their relative cellular concentrations, alterations in their subcellular location in response to other signalling pathways and regulation of their activity brought about by post-translational modifications. The binding properties of NCoR and SMRT are quite distinct from these cofactors since they contain extended LXXLL-like motifs (Hu & Lazar 1999) that cannot fit into the cleft formed on activated receptors but bind to certain nuclear receptors, including retinoic acid and thyroid hormone receptors, in the absence of hormone and to steroid receptors in the presence of antagonists (Huang et al. 2002, Liu et al. 2002, Shang & Brown 2002). Despite differences in the binding properties of cofactors to nuclear receptors they all seem to act as docking or scaffold proteins for the assembly of enzyme complexes at specific sites on a promoter. The function of these enzymes, which include acetylases/deacetylases, kinases/phosphatases and methyl transferases, is to catalyse the modification of histones that results in the remodelling of chromatin and ultimately gene activation or gene repression.
A schematic diagram of the structure of RIP140 is shown in Fig. 1![]() . It consists of 10 leucine-rich motifs that allow its recruitment to the ligand-binding domain of nuclear receptors (Heery et al. 1997; Torchia et al. 1997) and four repression domains (Christian et al. 2004). Individual LXXLL motifs are assumed to allow selective binding to different receptors; for example motif 10, LXXML, is reported to bind preferentially to retinoid receptors (Lee & Wei 1999). The repression domains exhibit autonomous activity suggesting that they function by recruiting enzymes that modify histones or DNA. Repression domain 1 has been found to bind histone deacetylases (HDACs) (Wei et al. 2000) while repression domain 2 binds C-terminal-binding protein (CtBP) that then binds enzymes, including histone deacetylases (Vo et al. 2001). The mechanism of action of repression domains 3 and 4 is unknown (Christian et al. 2004). Nevertheless it seems likely that the four repression domains allow the regulated recruitment of distinct sets of chromatin-modifying enzymes, each of which is capable of repressing transcription from target genes.
. It consists of 10 leucine-rich motifs that allow its recruitment to the ligand-binding domain of nuclear receptors (Heery et al. 1997; Torchia et al. 1997) and four repression domains (Christian et al. 2004). Individual LXXLL motifs are assumed to allow selective binding to different receptors; for example motif 10, LXXML, is reported to bind preferentially to retinoid receptors (Lee & Wei 1999). The repression domains exhibit autonomous activity suggesting that they function by recruiting enzymes that modify histones or DNA. Repression domain 1 has been found to bind histone deacetylases (HDACs) (Wei et al. 2000) while repression domain 2 binds C-terminal-binding protein (CtBP) that then binds enzymes, including histone deacetylases (Vo et al. 2001). The mechanism of action of repression domains 3 and 4 is unknown (Christian et al. 2004). Nevertheless it seems likely that the four repression domains allow the regulated recruitment of distinct sets of chromatin-modifying enzymes, each of which is capable of repressing transcription from target genes.
|
| Role of RIP140 in female reproduction |
|---|
|
|
|---|
|
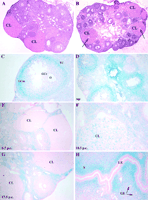
The precise site of RIP140 action in the ovary required for ovulation has not been determined definitively. RIP140 expression, as determined by analysing ß-galactosidase activity as a marker of promoter activity (Fig. 2![]() ), indicated that the highest levels are found in granulosa cells but low levels are also present in thecal and interstitial cells. No expression was detected in the granulosa cells of primary follicles or small follicles, but expression increased during follicular maturation, with the highest level detected in the outer mural cells in pre-ovulatory follicles (Fig. 2C
), indicated that the highest levels are found in granulosa cells but low levels are also present in thecal and interstitial cells. No expression was detected in the granulosa cells of primary follicles or small follicles, but expression increased during follicular maturation, with the highest level detected in the outer mural cells in pre-ovulatory follicles (Fig. 2C![]() ). Expression levels decreased dramatically after luteinization but were subsequently regulated in the corpora lutea of pregnant mice (Fig. 2D–G
). Expression levels decreased dramatically after luteinization but were subsequently regulated in the corpora lutea of pregnant mice (Fig. 2D–G![]() ). In ovaries of heterozygous or RIPKO mice used for embryo transfer there is negligible expression from the RIP140 promoter in corpora lutea at day 6.5 post-coitus (p.c.), but expression increases at mid-gestation, is maintained until at least day 13.5 p.c., and then declines to negligible levels at day 17.5 p.c., correlating with initiation of the regression of the corpora lutea. While initial failure of oocyte release did not affect luteinization and production of progesterone in the early stages of pregnancy, the normal mid-gestation rise in progesterone levels was not as pronounced in embryo-transferred RIP140-null mice. This is despite the fact that the corpora lutea were still large and apparently healthy (Leonardsson et al. 2002).
). In ovaries of heterozygous or RIPKO mice used for embryo transfer there is negligible expression from the RIP140 promoter in corpora lutea at day 6.5 post-coitus (p.c.), but expression increases at mid-gestation, is maintained until at least day 13.5 p.c., and then declines to negligible levels at day 17.5 p.c., correlating with initiation of the regression of the corpora lutea. While initial failure of oocyte release did not affect luteinization and production of progesterone in the early stages of pregnancy, the normal mid-gestation rise in progesterone levels was not as pronounced in embryo-transferred RIP140-null mice. This is despite the fact that the corpora lutea were still large and apparently healthy (Leonardsson et al. 2002).
RIP140 may also play a role in uterine function given that it is in the glandular epithelium, stroma and myometrium of the non-pregnant uterus (Fig. 2H![]() ). To investigate its function during pregnancy embryo-transfer experiments were performed. The transfer of wild-type embryos into pseudopregnant RIPKO females resulted in a similar number of implantation sites as in wild-type mice, indicating that uterine RIP140 is not required for implantation. Nevertheless RIP140 was expressed in primary decidual cells around the time of implantation and, by 9.5 days p.c., in differentiating decidual cells on the anti-mesometrial side of the uterus (Leonardsson et al. 2002). At later stages of pregnancy, however, there was a progressive reduction in the number of surviving embryos in the RIP140-null mice, suggesting that uterine RIP140 expression might be required to maintain the pregnancy. In addition, while the majority of pups born from control mothers survived after birth, 75% of the pups from RIP140-null mothers were dead within 24 h (Leonardsson et al. 2002).
). To investigate its function during pregnancy embryo-transfer experiments were performed. The transfer of wild-type embryos into pseudopregnant RIPKO females resulted in a similar number of implantation sites as in wild-type mice, indicating that uterine RIP140 is not required for implantation. Nevertheless RIP140 was expressed in primary decidual cells around the time of implantation and, by 9.5 days p.c., in differentiating decidual cells on the anti-mesometrial side of the uterus (Leonardsson et al. 2002). At later stages of pregnancy, however, there was a progressive reduction in the number of surviving embryos in the RIP140-null mice, suggesting that uterine RIP140 expression might be required to maintain the pregnancy. In addition, while the majority of pups born from control mothers survived after birth, 75% of the pups from RIP140-null mothers were dead within 24 h (Leonardsson et al. 2002).
The high incidence of fetal loss in mid-pregnancy and the death of the majority of pups at or shortly after birth following embryo transfer suggests that some aspects of the maternal expression of RIP140 are sub-optimal for the maintenance of pregnancy and for the subsequent survival of offspring. However, the ovarian-transfer experiments indicated that uterine and mammary gland RIP140 expression is not essential for pregnancy and survival. We conclude from this apparent disparity that the pregnancy failure and the death of pups may reflect a defect in the RIP140-null ovary which fails to support fully the function of the uterus and the mammary gland.
| Role of RIP140 in adipose tissue |
|---|
|
|
|---|
Further investigation showed that the RIP140-null mice are lean because they fail to store triglycerides and are resistant to obesity induced by high-fat diet, suggesting that alternative mechanisms are involved in the dissipation of excess fuel (Leonardsson et al. 2004). The most striking finding was the upregulation, by more than 100-fold, of mitochondrial uncoupling protein 1 (UCP1) mRNA, in inguinal WAT of RIPKO mice. In RIPKO mice fed on a normal diet, adipocyte volume was 2.7-fold less in inguinal WAT, compared with wild-type littermates, while there were no differences in the adipocytes in the interscapular brown adipose tissue (BAT), and there was no evidence from Oil Red O staining or MRI/MRS that fat was being stored in alternative tissues such as the liver. In animals fed on a high-fat diet RIPKO mice gained less weight than wild-type mice and showed a smaller increase in adipocyte volume. Lipid accumulation in the liver on high-fat diet was seen in wild-type but not in RIPKO mice, showing that the absence of RIP140 protects against hepatic steatosis (Leonardsson et al. 2004).
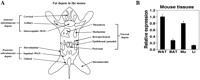
WAT and BAT can be distinguished by the morphology of the adipocytes; unilocular white adipocytes store energy as triglycerides and release it when required whereas multilocular brown adipocytes dissipate energy as heat. White and brown adipocytes are distributed in different proportions in distinct fat depots in mice, and the depots can be classified as subcutaneous or visceral in origin (Fig. 3A![]() ). In mature animals, most of the BAT is found in the interscapular depot, but in young animals it can also be found in the posterior subcutaneous (dorsolumbar, inguinal and gluteal), mediastinic and perirenal depots, mainly composed of WAT. Visceral WAT depots include mesenteric and gonadal, i.e. epididymal or ovarian, fat (Cinti 2002).
). In mature animals, most of the BAT is found in the interscapular depot, but in young animals it can also be found in the posterior subcutaneous (dorsolumbar, inguinal and gluteal), mediastinic and perirenal depots, mainly composed of WAT. Visceral WAT depots include mesenteric and gonadal, i.e. epididymal or ovarian, fat (Cinti 2002).
|
|
 ) controls fat accumulation in subcutaneous rather than visceral adipose tissue in females by upregulating 2A-adrenergic receptors (Pedersen et al. 2004). Such depot-specific expression of key genes involved in fat deposition and mobilization may also be mediated by peroxisome proliferator-activated receptor-(PPAR-), since PPAR- agonists, used to treat insulin resistance, exert depot-specific effects upon human fat deposition (Laplante et al. 2003). In rats, leptin mRNA levels are higher in gonadal and retroperitoneal (intra-abdominal depots) than in inguinal (subcutaneous) adipose tissues (Zheng et al. 1996), while glucocorticoid receptor expression is higher in epididymal than inguinal and retro-peritoneal depots (Zhang et al. 2002). Angiotensinogen expression changes on high-fat feeding in visceral fat depots in mice, but not in subcutaneous depots nor in BAT (Rahmouni et al. 2004). The differential regulation of adipocyte function between subcutaneous and visceral depots in humans is an important area for investigation because increased visceral adipose tissue in human obesity is associated with pathological effects such as hypertension and metabolic syndrome, whereas subcutaneous fat deposition is less harmful.
) controls fat accumulation in subcutaneous rather than visceral adipose tissue in females by upregulating 2A-adrenergic receptors (Pedersen et al. 2004). Such depot-specific expression of key genes involved in fat deposition and mobilization may also be mediated by peroxisome proliferator-activated receptor-(PPAR-), since PPAR- agonists, used to treat insulin resistance, exert depot-specific effects upon human fat deposition (Laplante et al. 2003). In rats, leptin mRNA levels are higher in gonadal and retroperitoneal (intra-abdominal depots) than in inguinal (subcutaneous) adipose tissues (Zheng et al. 1996), while glucocorticoid receptor expression is higher in epididymal than inguinal and retro-peritoneal depots (Zhang et al. 2002). Angiotensinogen expression changes on high-fat feeding in visceral fat depots in mice, but not in subcutaneous depots nor in BAT (Rahmouni et al. 2004). The differential regulation of adipocyte function between subcutaneous and visceral depots in humans is an important area for investigation because increased visceral adipose tissue in human obesity is associated with pathological effects such as hypertension and metabolic syndrome, whereas subcutaneous fat deposition is less harmful. | Other sites of RIP140 action |
|---|
|
|
|---|
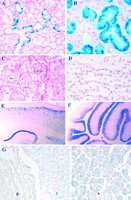
Expression of RIP140, as revealed by detection of ß-galactosidase by histochemical staining, showed that the gene is expressed in specific cells in a variety of other tissues, mainly in hormone-responsive cell types. In the salivary gland the gene is expressed in ducts of the sublingual gland and in granular convoluted tubules of the submandibular gland (Fig. 5A and B![]() ). The differentiation and maintenance of granular convoluted tubule cells are under the control of the synergistic actions of androgens, thyroid hormones and adrenocortical hormones (Kim et al. 2001), suggesting that nuclear receptor cofactors have a role to play in these cells. In the kidney RIP140 is expressed in a variety of epithelial cells, in proximal and distal convoluted tubules, the loop of Henle and collecting ducts (Fig. 5C and D
). The differentiation and maintenance of granular convoluted tubule cells are under the control of the synergistic actions of androgens, thyroid hormones and adrenocortical hormones (Kim et al. 2001), suggesting that nuclear receptor cofactors have a role to play in these cells. In the kidney RIP140 is expressed in a variety of epithelial cells, in proximal and distal convoluted tubules, the loop of Henle and collecting ducts (Fig. 5C and D![]() ). In male mice RIP140 is expressed in epididymal epithelial cells, in the prostate and the testis. In addition, ß-galactosidase staining is present in blood vessels and neuronal ganglia in a variety of tissues.
). In male mice RIP140 is expressed in epididymal epithelial cells, in the prostate and the testis. In addition, ß-galactosidase staining is present in blood vessels and neuronal ganglia in a variety of tissues.
|
| Conclusion |
|---|
|
|
|---|
RIP140 may therefore provide a novel therapeutic target for the treatment of obesity and related disorders. Further investigation of the role of this corepressor in ovulation may bring about new opportunities for treating infertility or the development of new contraceptive strategies.
| References |
|---|
|
|
|---|
Christian M, Tullet JM & Parker MG 2004 Characterization of four autonomous repression domains in the corepressor receptor interacting protein 140. Journal of Biological Chemistry 279 15645–15651.
Cinti S 2002 Adipocyte differentiation and transdifferentiation: plasticity of the adipose organ. Journal of Endocrinological Investigation 25 823–835.[Web of Science][Medline]
Heery DM, Kalkhoven E, Hoare S & Parker MG 1997 A signature motif in transcriptional co-activators mediates binding to nuclear receptors. Nature 387 733–736.[CrossRef][Medline]
Hu X & Lazar MA 1999 The CoRNR motif controls the recruitment of corepressors by nuclear hormone receptors. Nature 402 93–96.[CrossRef][Medline]
Huang HJ, Norris JD & McDonnell DP 2002 Identification of a negative regulatory surface within estrogen receptor alpha provides evidence in support of a role for corepressors in regulating cellular responses to agonists and antagonists. Molecular Endocrinology 16 1778–1792.
Jepsen K, Hermanson O, Onami TM, Gleiberman AS, Lunyak V, McEvilly RJ, Kurokawa R, Kumar V, Liu F, Seto E et al. 2000 Combinatorial roles of the nuclear receptor corepressor in transcription and development. Cell 102 753–763.[CrossRef][Web of Science][Medline]
Kim J, Amano O, Wakayama T, Takahagi H & Iseki S 2001 The role of cyclic AMP response element-binding protein in testosterone-induced differentiation of granular convoluted tubule cells in the rat submandibular gland. Archives of Oral Biology 46 495–507.[CrossRef][Web of Science][Medline]
Laplante M, Sell H, MacNaul KL, Richard D, Berger JP & Deshaies Y 2003 PPAR-gamma activation mediates adipose depot-specific effects on gene expression and lipoprotein lipase activity: mechanisms for modulation of postprandial lipemia and differential adipose accretion. Diabetes 52 291–299.
Lee CH & Wei LN 1999 Characterization of receptor-interacting protein 140 in retinoid receptor activities. Journal of Biological Chemistry 274 31320–31326.
Leonardsson G, Jacobs MA, White R, Jeffery R, Poulsom R, Milligan S & Parker M 2002 Embryo transfer experiments and ovarian transplantation identify the ovary as the only site in which nuclear receptor interacting protein 1/RIP140 action is crucial for female fertility. Endocrinology 143 700–707.
Leonardsson G, Steel JH, Christian M, Pocock V, Milligan S, Bell J, So PW, Medina-Gomez G, Vidal-Puig A, White R et al. 2004 Nuclear receptor corepressor RIP140 regulates fat accumulation. PNAS 101 8437–8442.
L’Horset F, Dauvois S, Heery DM, Cavailles V & Parker MG 1996 RIP-140 interacts with multiple nuclear receptors by means of two distinct sites. Molecular and Cellular Biology 16 6029–6036.
Liu Z, Auboeuf D, Wong J, Chen JD, Tsai SY, Tsai MJ & O’Malley BW 2002 Coactivator/corepressor ratios modulate PR-mediated transcription by the selective receptor modulator RU486. PNAS 99 7940–7944.
McKenna NJ & O’Malley BW 2002 Combinatorial control of gene expression by nuclear receptors and coregulators. Cell 108 465–474.[CrossRef][Web of Science][Medline]
Nagy L, Kao HY, Chakravarti D, Lin RJ, Hassig CA, Ayer DE, Schreiber SL & Evans RM 1997 Nuclear receptor repression mediated by a complex containing SMRT, mSin3A, and histone deacetylase. Cell 89 373–380.[CrossRef][Web of Science][Medline]
Pedersen SB, Kristensen K, Hermann PA, Katzenellenbogen JA & Richelsen B 2004 Estrogen controls lipolysis by up-regulating alpha2A-adrenergic receptors directly in human adipose tissue through the estrogen receptor alpha. Implications for the female fat distribution. Journal of Clinical Endocrinology and Metabolism 89 1869–1878.
Rahmouni K, Mark AL, Haynes WG & Sigmund CD 2004 Adipose depot-specific modulation of angiotensinogen gene expression in diet-induced obesity. American Journal of Physiology – Endocrinology and Metabolism 286 E891–E895.
Rossmeisl M, Barbatelli G, Flachs P, Brauner P, Zingaretti MC, Marelli M, Janovska P, Horakova M, Syrovy I, Cinti S et al. 2002 Expression of the uncoupling protein 1 from the aP2 gene promoter stimulates mitochondrial biogenesis in unilocular adipocytes in vivo. European Journal of Biochemistry 269 19–28.[Web of Science][Medline]
Shang Y & Brown M 2002 Molecular determinants for the tissue specificity of SERMs. Science 295 2465–2468.
Spiegelman BM & Flier JS 2001 Obesity and the regulation of energy balance. Cell 104 531–543.[CrossRef][Web of Science][Medline]
Subramaniam N, Treuter E & Okret S 1999 Receptor interacting protein RIP140 inhibits both positive and negative gene regulation by glucocorticoids. Journal of Biological Chemistry 274 18121–18127.
Torchia J, Rose DW, Inostroza J, Kamei Y, Westin S, Glass CK & Rosenfeld MG 1997 The transcriptional co-activator p/CIP binds CBP and mediates nuclear-receptor function. Nature 387 677–684.[CrossRef][Medline]
Vo N, Fjeld C & Goodman RH 2001 Acetylation of nuclear hormone receptor-interacting protein RIP140 regulates binding of the transcriptional corepressor CtBP. Molecular and Cellular Biology 21 6181–6188.
Wei LN, Hu X, Chandra D, Seto E & Farooqui M 2000 Receptor-interacting protein 140 directly recruits histone deacetylases for gene silencing. Journal of Biological Chemistry 275 40782–40787.
White R, Leonardsson G, Rosewell I, Ann Jacobs M, Milligan S & Parker M 2000 The nuclear receptor co-repressor nrip1 (RIP140) is essential for female fertility. Nature Medicine 6 1368–1374.[CrossRef][Web of Science][Medline]
Zhang Y, Guo KY, Diaz PA, Heo M & Leibel RL 2002 Determinants of leptin gene expression in fat depots of lean mice. American Journal of Physiology – Regulatory, Integrative and Comparative Physiology 282 R226–R234.
Zheng D, Jones JP, Usala SJ & Dohm GL 1996 Differential expression of ob mRNA in rat adipose tissues in response to insulin. Biochemical and Biophysical Research Communications 218 434–437.[CrossRef][Web of Science][Medline]
Received 6 July 2004
Accepted 2 September 2004
This article has been cited by other articles:
 |
I. Zschiedrich, U. Hardeland, A. Krones-Herzig, M. Berriel Diaz, A. Vegiopoulos, J. Muggenburg, D. Sombroek, T. G. Hofmann, R. Zawatzky, X. Yu, et al. Coactivator function of RIP140 for NF{kappa}B/RelA-dependent cytokine gene expression Blood, July 15, 2008; 112(2): 264 - 276. [Abstract] [Full Text] [PDF] |
||||
|
|
R. Sanchez-Martinez, A. Zambrano, A. I. Castillo, and A. Aranda Vitamin D-Dependent Recruitment of Corepressors to Vitamin D/Retinoid X Receptor Heterodimers Mol. Cell. Biol., June 1, 2008; 28(11): 3817 - 3829. [Abstract] [Full Text] [PDF] |
||||
|
|
M. M. Rytinki and J. J. Palvimo SUMOylation Modulates the Transcription Repressor Function of RIP140 J. Biol. Chem., April 25, 2008; 283(17): 11586 - 11595. [Abstract] [Full Text] [PDF] |
||||
|
|
R. Bartz, J. Seemann, J. K. Zehmer, G. Serrero, K. D. Chapman, R. G.W. Anderson, and P. Liu Evidence that Mono-ADP-Ribosylation of CtBP1/BARS Regulates Lipid Storage Mol. Biol. Cell, August 1, 2007; 18(8): 3015 - 3025. [Abstract] [Full Text] [PDF] |
||||
|
|
P. Gupta, M. D. M. Huq, S. A. Khan, N.-P. Tsai, and L.-N. Wei Regulation of Co-repressive Activity of and HDAC Recruitment to RIP140 by Site-specific Phosphorylation Mol. Cell. Proteomics, November 1, 2005; 4(11): 1776 - 1784. [Abstract] [Full Text] [PDF] |
||||
|
|
J. M. A. Tullet, V. Pocock, J. H. Steel, R. White, S. Milligan, and M. G. Parker Multiple Signaling Defects in the Absence of RIP140 Impair Both Cumulus Expansion and Follicle Rupture Endocrinology, September 1, 2005; 146(9): 4127 - 4137. [Abstract] [Full Text] [PDF] |
||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| HOME | HELP | CONTACT US | SUBSCRIPTIONS | ARCHIVE | SEARCH | TABLE OF CONTENTS |